
From Sensory to Long-Term Memory
Evidence from Auditory Memory Reactivation Studies
Istva
´
n Winkler
1,2
and Nelson Cowan
3
1
Institute for Psychology, Hungarian Academy of Sciences, Hungary,
2
Cognitive Brain Research Unit, Department of Psychology, University of Helsinki, Finland,
3
Department of Psychological Sciences, University of Missouri, USA
Abstract. Everyday experience tells us that some types of auditory sensory information are retained for long periods of
time. For example, we are able to recognize friends by their voice alone or identify the source of familiar noises even years
after we last heard the sounds. It is thus somewhat surprising that the results of most studies of auditory sensory memory
show that acoustic details, such as the pitch of a tone, fade from memory in ca. 10Ð15 s. One should, therefore, ask (1) what
types of acoustic information can be retained for a longer term, (2) what circumstances allow or help the formation of
durable memory records for acoustic details, and (3) how such memory records can be accessed. The present review discusses
the results of experiments that used a model of auditory recognition, the auditory memory reactivation paradigm. Results
obtained with this paradigm suggest that the brain stores features of individual sounds embedded within representations of
acoustic regularities that have been detected for the sound patterns and sequences in which the sounds appeared. Thus, sounds
closely linked with their auditory context are more likely to be remembered. The representations of acoustic regularities are
automatically activated by matching sounds, enabling object recognition.
Keywords: memory, auditory sensory memory, long-term sensory memory, reactivation, event-related brain potentials, mis-
match negativity (MMN)
1. Introduction
Traditionally, the processing of sensory and categori-
cal information have been distinguished from each
other on the basis of performance differences found
in some experimental procedures and by evidence
showing anatomical separation of sensory and cate-
gorical processing in the human brain (see, however,
Näätänen, Tervaniemi, Sussman, Paavilainen, & Win-
kler, 2001, for recent evidence concerning the “intelli-
gent” functions of auditory sensory-specific areas in
the human brain). Previous research in experimental
psychology identified four important features charac-
terizing sensory memory traces, distinguishing them
from categorical memory representations (Broadbent,
1958; Cowan, 1984, 1988; for a full discussion of the
supporting and contradictory evidence, see Section
3.2.): 1) the formation of sensory memory traces does
not depend on attention; 2) the information stored in
sensory memory traces is modality-specific; and 3)
has a resolution, which is finer than the conventional
meaningful categories; but 4) it is lost within a short
period of time. The goal of the current review is to
reexamine these distinguishing features of sensory
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
DOI: 10.1027/1618-3169.52.1.3
memory traces in the light of recent evidence obtained
with electrophysiological and behavioral methods,
mostly using memory-reactivation procedures.
1.1 Properties of Sensory Memory:
Behavioral Studies
Classical multi-store models of memory postulated
separate stores for the retention of sensory informa-
tion (e.g., Atkinson & Shiffrin, 1968). The sensory
memory stores (a separate one for each modality)
were assumed to serve as temporary buffers from
which information could be accessed for a short time,
after which they were lost due to decay or to interfer-
ence from more recent stimuli. The information se-
lected from the sensory buffers was categorized, or
transformed into a common internal code, allowing
modality-independent operations. Only categorized
information was assumed to be stored in more durable
stores.
These features of the multi-store models corre-
spond well with the majority of the results of sensory
memory research. For example, it has been found in

4 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
many studies that subjects can only tell the difference
between two closely similar sounds if the sounds to
be compared are presented within ca. 10 s (taken to
suggest decay) Cowan, 1984). Also, presenting irrele-
vant sounds between the to-be-compared ones deterio-
rates performance, the more so, the closer the sim-
ilarity between the to-be-compared and the interven-
ing sounds (taken to suggest interference) (Cowan,
1984; Deutsch, 1975; Massaro, 1970). When the
sounds to be compared are separated by long silent
intervals, subjects can only discriminate them if they
substantially differ from each other. The degradation
of sensory resolution is compatible with the idea that
once the trace of the first sound is eliminated from
the sensory buffer, subjects can only rely on those
memory stores that have a longer retention interval.
These stores, however, only contain categorized infor-
mation representing stimulus features with a cruder
resolution than the sensory buffer. Importantly, it ap-
pears that the retention interval of the sensory buffer
cannot be extended much by top-down control (e.g.,
Keller, Cowan, & Saults, 1995), though there is a
small effect of that nature. Three related findings in
the area of speech perception leading toward the no-
tion of separated sensory and categorical memory
stores are (1) the category boundary effect, or more
successful discrimination between two phonemes fall-
ing across a category boundary than between two allo-
phones (instances of a single category) of comparable
physical separation, (2) the delay effect, or poorer
comparison of two allophones as a function of the
temporal separation between these allophones in the
range of a few seconds, and (3) the vowel advantage,
or much more rapid forgetting of allophonic detail for
the acoustically-complex stop consonants than for the
acoustically-simpler vowels (Fujisaki & Kawashima,
1971; Pisoni, 1973). Acoustic theory has emphasized
this distinction between unstable sensory information
and more stable categorical information (e.g., Dur-
lach & Braida, 1969).
However, there also exist results indicating that not
all sensory information is lost within a few seconds.
Craik and Kirsner (1974) reviewed studies showing
that people often remember voice information for
longer than the period of 30 s or so that has been the
presumed duration of auditory sensory memory, and
they carried out their own interesting experiments re-
inforcing that point. In their studies, spoken target
words were presented 4 s apart in two different voices
(male and female) in random order. The test for each
word consisted of another version of the word (the
probe word) that had to be recognized. Foils that had
not been presented as targets also appeared as negative
probes, and the question was whether the probe had
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
appeared before in the experiment. The distance be-
tween the target and the corresponding probe word was
a lag of 1, 2, 4, 8, 16, or 32 words. At a lag of 1, the
target and probe occurred with no intervening word; at
a lag of 2, they occurred with one intervening word; and
so on. It was found in several of the experiments that
spoken probe words were recognized faster and more
accurately when the voice remained the same from the
target word to the probe word. This occurred even at the
long delays, indicating that some memory of the voice
persisted for over 2 min in the presence of intervening
words. It also was found that subjects maintained an
ability to recall (at better-than-chance accuracy) the
voice in which a particular word had been presented.
For example, in one experiment, in which the probe
words were visually presented, the proportion of recall
of the voice conditional upon correct recognition of the
word was, at the six lags, 1.00, 0.98, 0.87, 0.75, 0.76,
and 0.73, respectively. Thus, at lags of 8 words (32 s)
and above, recall of the voice reached an asymptote of
much-better-than-chance accuracy.
In a very different type of procedure demonstrating
long-term storage of memory for sound, Crowder
(1989) presented a pure tone followed by a note played
by a musical instrument. Comparison of the pitches of
the two sounds was speeded when participants had ad-
vanced knowledge of which instrument was to be used
for the second sound. Presumably, long-term memory
was used to generate a mental image of the frequency
of the pure tone as played in the timbre of the instru-
ment that was used for the second sound.
As a consequence, modern memory models sug-
gest that sensory stimulus codes are processed along
with categorical stimulus representations by special-
ized subsystems or activation processes (e.g., see
Cowan, 1988, 1995, 1999). However, although these
models assume that some sensory details are retained
for longer periods of time, they do not explain the
apparent contradiction between the classical findings
of limitations in accessing sensory information and
longer-term retention of these data. Our approach to
this issue was based on everyday experience. One can
recognize concrete objects (not just object categories)
by sensory details even long after the object was last
encountered. For example, we are often able to recog-
nize our friends’ voices over the telephone or shouting
from another room, even though there are no known
categorical properties distinguishing one voice from
another. A model of the recognition situation may re-
veal important information about what type of sensory
information is retained in the brain as well as about
the circumstances that help in forming such memory
representations. The auditory memory reactivation
paradigm was designed to test these questions. By ba-

5I. Winkler & N. Cowan: From Sensory to Long-Term Memory
sing the test of “recognition” on an event-related brain
potential (ERP) that can be measured independently
of the subject’s task, we avoided confounding factors
stemming from the task and strategy of the subject,
thus providing an unbiased measure of what sensory
information is stored in the brain for longer time
periods.
1.2 The Mismatch Negativity
Event-Related Potential
Following a brief introduction to the ERP component
used to test the recognition of sounds, the reactivation
paradigm will be described in detail. We will show
that with this paradigm, we are able to tap auditory
sensory information that is resistant to decay and in-
terference and discuss what processes and memory
structures may underlie the reactivation phenomenon.
Evidence will be provided showing that reactivated
sound information can also be accessed in active
discrimination tasks. In the discussion, we turn to the
question of what kind of sound information is retained
for longer periods of time. Finally, we relate our find-
ings to two current models of working memory.
The ERP component involved in testing memory
reactivation has been termed the mismatch negativity
(MMN). MMN is elicited whenever a sound violates
some regular aspect of the preceding sound sequence
(for recent reviews, see Näätänen & Winkler, 1999;
Picton, Alain, Otten, & Ritter, 2000). The elicitation
of MMN requires the presence of some representation
of the violated acoustic regularity. The auditory sen-
sory information encoded in these regularity represen-
tations corresponds to that appearing in perception.
MMN is elicited whether or not the subject’s task is
related to the test sounds. In fact, in the majority of
MMN studies, subjects were engaged in some activity
involving visual stimuli (e.g., they read a book,
watched a movie, or performed some reaction task
with visual stimuli) and were instructed to disregard
the sounds presented to them (the “passive” condi-
tion). Thus MMN can be used to obtain a task-inde-
pendent index of the retention of auditory information
in the brain.
The MMN component typically peaks between 100
and 200ms from the onset of the regularity violation
with negative polarity over the fronto-central scalp and
positive over scalp locations above the opposite side of
the Sylvian fissure. This is because the main cortical
generators of MMN lie within or in the vicinity of the
supratemporal plane with additional contribution from
sources located in the frontal cortex (Halgren et al.,
1995; Opitz, Rinne, Mecklinger, von Cramon, & Schrö-
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
ger, 2002). The simplest and most commonly used
paradigm for obtaining MMN is the auditory oddball
sequence. When a repeating sound (termed the “stan-
dard” sound) is occasionally exchanged for a different
sound (“deviant” sound) MMN is elicited. However,
MMN is also elicited by violations of far more com-
plex auditory regularities (for reviews, see Näätänen et
al., 2001; Winkler, 2003). The MMN wave can be delin-
eated from other concurrent ERP components by
subtracting from the ERP response elicited by the devi-
ant stimulus the ERP elicited by some control sound.
For a good assessment of the MMN response, the con-
trol sound should share as many features as possible
with the deviant sound, but it should not itself elicit an
MMN (i.e., it should be a stimulus that conforms to the
regularities of the sound sequence in which it appears).
The current explanation of MMN elicitation sug-
gests that incoming sounds are compared with extrap-
olations (sensory inferences) calculated from the rep-
resentation of regularities detected in the preceding
sound sequence. Sounds that mismatch these extrapo-
lations activate the MMN-generating process (Win-
kler, Karmos, & Näätänen, 1996b). It is important to
note that the presence of a sensory memory record of
a sound in the brain is not a sufficient prerequisite of
MMN elicitation; MMN is only elicited once some
auditory regularity has been detected and a subsequent
sound violates this regularity (Cowan, Winkler,
Teder, & Näätänen, 1993; Sussman, Sheridan, Kreu-
zer, & Winkler, 2003a; Winkler, Schröger, & Cowan,
2001). Thus, stimulus change per se (e.g., two dif-
ferent sounds presented successively at the beginning
of a sound sequence or within an ever-changing se-
quence of sounds) does not result in MMN elicitation
(Cowan et al., 1993; Horva
´
th, Czigler, Sussman, &
Winkler, 2001; Winkler, 1996).
The MMN-generating process appears to be inde-
pendent of top-down control (Rinne, Antila, & Wink-
ler, 2001; Sussman, Winkler, & Wang, 2003b). It has
been shown that in most cases, the MMN results ob-
tained in the “passive” situation match those that can
be obtained in the same paradigm with attention di-
rected away from the sounds in a controlled manner
(Sussman et al., 2003b; Winkler et al., 2003). How-
ever, it should be noted that some of the processes
underlying the detection of auditory regularities can
be modulated by top-down control and the outcome
of these processes is also reflected in the elicitation
and/or amplitude of the MMN response (Sussman,
Winkler, Huotilainen, Ritter, & Näätänen, 2002; Suss-
man et al., 2003b). The possible functions of the
MMN-generating process are to initiate further pro-
cessing of the deviant sounds (Nääatänen, 1990) and/
or to update the regularity representations that did not
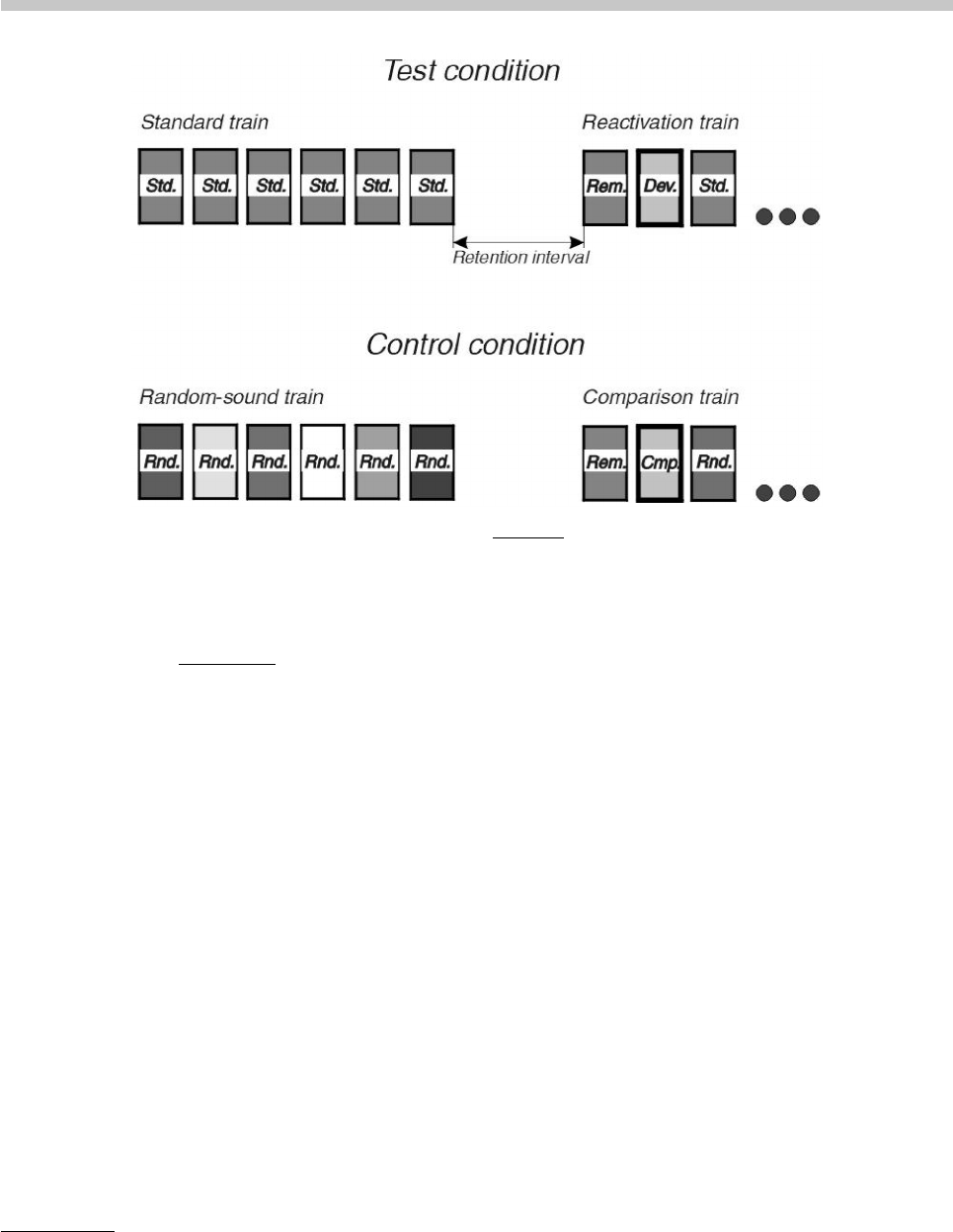
6 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Figure 1. Schematic illustration of an MMN reactivation paradigm. Top panel: The test condition. Standard trains consist of sounds
or sound patterns, which conform to some regularity. Elements of these trains are termed “standards”, marked with “Std” on the
figure. Standard trains are followed by the retention interval and the reactivation train. The first element of the reactivation train is
termed the “reminder” (marked “Rem”). It conforms to the regularities of the standard train (i.e., it is also a “standard”). Reactivation
is tested by the second element of the reactivation train, which violates some regularity of the standard train. Thus it is termed
“deviant”, marked as “Dev”. The reactivation train ends with a further standard or standards (their role is to provide a homogeneous
context for the deviant). Bottom panel: One possible control condition. Standard trains are exchanged for trains in which the sounds
or sound patterns vary in the regular feature of the standard trains (e.g., if the regularity used in the test condition is the constancy
of tone frequency, then frequency is randomly varied in the control condition). The trains substituted for the standard trains are
termed random-sound trains and their elements marked with “Rnd”. Reactivation trains are exchanged for comparison trains that
start with the same sounds as the reactivation trains (the reminder and a deviant, termed comparison tone and marked as “Cmp”).
The rest of the train is made up of random sounds, again varying in the critical regularity of the test condition.
correctly predict the deviant (Winkler & Czigler,
1998; Winkler et al., 1996b).
2. Auditory Memory Reactivation
2.1. The Reactivation Paradigm
For testing reactivation with the MMN measure, at
least two trains of sounds are needed (Figure 1, top
panel). The first train sets up a regularity (tone repeti-
tion in Figure 1). It is termed the “standard train.” The
standard train is followed by the retention interval.
The second train (“reactivation train”) starts with a
sound (or sound pattern) that conforms to the regulari-
ties of the standard train. This sound is termed the
1
The requirement of establishing a regularity in the standard train provides a good possibility for delineating the MMN
component from other overlapping ERP responses. Figure 1 (bottom panel) shows the optimal control sequence. Standard trains are
exchanged for trains that do not show the same regularity (termed “random-sound trains”). For example, if the deviant is set up to
violate the common frequency of the standard-train tones, then a train of tones randomly varying in frequency can be used in the
control condition. The “reminder” and the “deviant” are unchanged (compared with the test condition) but are again followed by
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
“reminder.” The second sound of the reactivation train
then violates the regularity of the standard train. This
sound is termed the deviant or position-2 deviant in
paradigms testing MMN elicitation also in later posi-
tions of the trains. In some paradigms, each train
served both functions: the first two sounds of the train
tested reactivation with respect to the preceding train,
whereas later sounds in the train set up the standard
for the next train. If the (position-2) deviant sound
elicits the MMN component, one can conclude that
the regularity of the standard train was represented in
the brain and this representation was available to the
MMN-generating process. This is because, as was
mentioned in the previous section, stimulus change
alone does not activate the MMN-generating process.
MMN can only be elicited if the deviant sound vio-
lates some detected regularity.
1
Figure 2 (top left
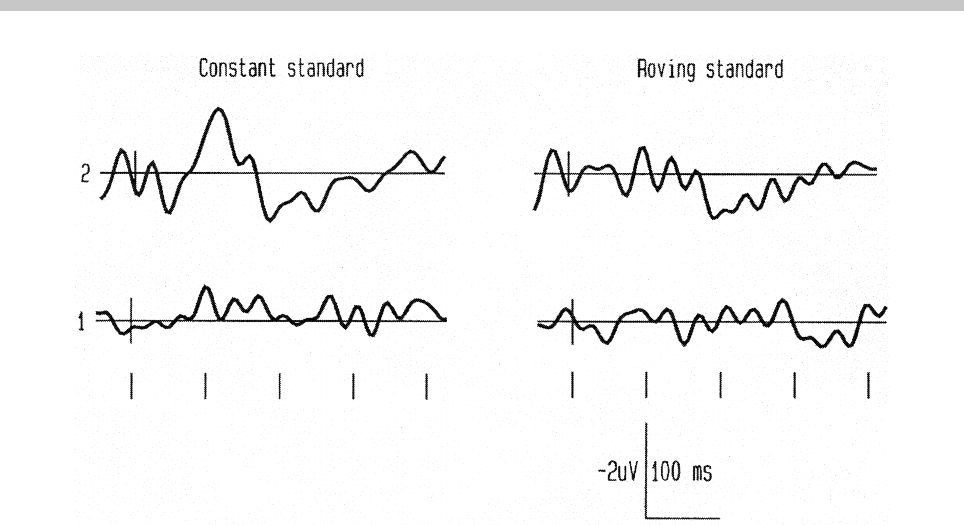
7I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Figure 2. ERP difference waves (deviant minus same-position standard-tone responses) obtained by Cowan et al. (1993) at the
frontal (Fz) electrode location in position 2 (reactivation, first row) and in position 1 (deactivation test, second row) of the tone
trains. In the constant standard condition (left column), standard and deviant tones were fixed within the stimulus blocks. In the
roving standard condition (right column), the frequency of both standard and deviant tones changed from train to train. Tone onset
is at the crossing of the x and y axes. Calibration is marked at the lower right corner. Tick-marks at the bottom are spaced 100ms
apart. The MMN response, which appears only in position 2 of the constant standard condition (upper left corner), is a negative
wave peaking between 100and 200ms post-stimulus.
panel; adapted from Cowan et al., 1993) shows the
MMN response elicited by a position-2 frequency-de-
viant tone.
However, the elicitation of MMN by a position-2 de-
viant, in and of itself, does not prove that reactivation
occurred. One should also check whether the represen-
tation of the regularity of the standard train was still
available to the MMN-generating process following the
retention interval. That is, it should be tested whether a
deviant presented in the first position of a stimulus train
elicits the MMN. Figure 2 (bottom panels) shows that
in Cowan et al.’s (1993) study no MMN was elicited by
position-1 deviant tones. It has been repeatedly found
that when trains are separated by a silent interval of 11 s
or more, no MMN is elicited by deviant sounds pre-
sented at the beginning of a train (Cowan et al., 1993;
Gaeta, Friedman, Ritter, & Cheng, 2001; Winkler et al.,
2001). Therefore, when the retention interval is longer
than 11 s, MMN elicited by position-2 deviants tells
that the representation of the regularity of the standard
train, which was not available to the MMN-generating
process at the beginning of the reactivation train be-
random sounds. This setup ensures that the control-condition deviant will not elicit MMN while the rest of its ERP components
match those elicited in the test condition. The first reactivation experiments used for comparison the response elicited by a standard
sound that was presented in the same position as the deviant. More recent studies used the control described above.
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
came available by the time the second sound of the reac-
tivation train was processed. Cowan et al. (1993)
termed this phenomenon “memory reactivation” as
they regarded it to be similar to the reactivation phe-
nomena described by Rovee-Collier and Hayne (1987).
Note, however, that the findings of Rovee-Collier and
Hayne refer to learned actions rather than sensory infor-
mation and that the timescale of their effect is much
longer (several days) than that ever tested with the cur-
rent reactivation paradigm.
The reminder is a critical element of the reactiva-
tion paradigm. Cowan et al. (1993) found that when
the first tone of a train substantially differed from the
tone repeated in the preceding train, it did not set up
a subsequent “deviant” for MMN elicitation. In their
“roving-standard” condition, Cowan et al. presented
short trains of tones in which all but possibly one tone
had the same (standard) frequency. This standard fre-
quency, however, changed from train to train. In ca.
17 % of the trains, a tone whose frequency differed
from the current as well as from the previous standard
frequency (deviant) appeared in the second position

8 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
of the train. The first tone of the train had the fre-
quency of the new standard, which was different from
that of the previous train. Position-2 deviants did not
elicit MMN in this situation (Figure 2, top right
panel). One could still argue that the roving-standard
condition of Cowan et al. did not set up a stable regu-
larity, because the standard frequency changed from
train to train. However, Ritter and his colleagues (Rit-
ter, Sussman, Molholm, & Foxe, 2002) have shown
that reactivation occurs even if the standard frequency
changes from train to train, when the reminder
matches the standard of the preceding train. Winkler
et al. (2002) narrowed the frequency range within
which the reminder is effective (i.e., reactivation oc-
curs). These authors found no MMN elicited by devi-
ants following a reminder that differed only by 3 %
from the standard frequency. Winkler et al.’s result
suggests that the representation against which the re-
minder is checked encodes features of the standard
sounds with a resolution characteristic of sensory
memory traces. Thus reactivation can indicate the
existence of finely resolved auditory information in
the human brain.
2.2. Characteristics of the Memory
Involved in Reactivation
The first question is whether the characteristics the
memory traces involved in reactivation are the same
as those describing the classical notion of auditory
sensory memory. Auditory sensory memory is subject
both to decay (estimates ranging from 10 to 20 s) and
to interference by similar sounds (Cowan, 1984). The
first reactivation study (Cowan et al., 1993) tested re-
tention intervals just beyond the most commonly ac-
cepted value for the duration of auditory sensory
memory (11Ð15 s). It is thus possible that a weak resi-
due of the auditory sensory memory trace of the re-
peated standard tone could have been reinforced by
the reminder. However, in a more recent study (Wink-
ler et al., 2002), the retention interval was set to 30 s.
The standard was again a repeating tone and the devi-
ant tone differed from it in frequency. All subjects
showed reactivation in this situation, even though only
one of them performed above chance level in discrimi-
nating the standard and deviant tones when the tones
were separated by a 30-s silent interval.
Interference from similar sounds has also been
tested (Winkler, Cowan, Cse
´
pe, Czigler, & Näätänen,
1996). Trains were composed of 12 tones. At least 5
of the first 6 tones were identical, whereas tones 7
through 12 (termed intervening tones) varied ran-
domly in frequency. Deviants appearing within the
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
first 6 tones also differed from the standard tones in
frequency. Trains were separated by 5.9 s of silence.
No MMN was elicited by the random-frequency in-
tervening tones presented in the last 3 positions of the
train (from position 10Ð12, i.e., the 4thÐ6th interven-
ing tones). Because these tones deviated from the
standard in the same way as the deviant did, the fact
that no MMN was elicited by them at the end of the
trains demonstrated that the frequency-regularity rep-
resentation was no longer available to the MMN-gen-
erating process after the 1st through 3rd intervening
tones, even before the onset of the silent interval that
separated the trains. Therefore, if the sensory informa-
tion involved in reactivation was vulnerable to inter-
ference from similar sounds, no MMN should be ex-
pected to be elicited by position-2 deviants (i.e., the
deviants testing reactivation following the silent in-
terval separating the trains). However, MMN was elic-
ited by position-2 deviants in this situation. This result
suggests that the memory involved in reactivation is
not subject to the type of sensory interference that
characterizes short-term auditory sensory memory.
The experiments reviewed above set up tone repeti-
tion as the standard. In most natural situations, however,
sound sequences include substantial amount of vari-
ance and the regularities have to be extracted from the
ever-changing input. One possible way to model such
variability is to set random changes in some sound
features while fixing the level of others. Ritter,
Gomes, Cowan, Sussman, and Vaughan (1998)
showed that constant tone-intensity was reactivated
when tone-frequency was varied throughout the stimu-
lus trains (including the frequency of the reminder
tone) and vice versa, constant tone-frequency was re-
activated when tone-intensity was varied. These re-
sults suggest that the memory representation involved
in reactivation encodes constancies extracted from the
variable input.
One could, however, argue that feature constancies
may be detected directly by neurons sensitive to nar-
row ranges of auditory features (see, e.g., Ritter, Dea-
con, Gomes, Javitt, & Vaughan, 1995). Furthermore,
the type of sensory information whose recognition we
intended to model with the reactivation paradigm is
based on secondary, rather than primary sound fea-
tures. For example, the timbre of a human voice that
allows one to recognize a friend by his/her voice alone
is characterized by the ratio between energy emitted
in certain frequency ranges, rather than by the abso-
lute pitch of the voice. A simple model of a secondary
auditory feature has been constructed by Saarinen,
Paavilainen, Schröger, Tervaniemi, and Näätänen
(1992), who presented tone pairs, 90% of which were
ascending in frequency (i.e., the frequency of the se-

9I. Winkler & N. Cowan: From Sensory to Long-Term Memory
cond tone of the pair was higher than that of the first
tone). The absolute frequencies of the tones varied
randomly throughout the sequences. Infrequent de-
scending-pitched tone pairs and tone repetitions elic-
ited the MMN response (for a study controlling all
aspects of pitch ascension, see Paavilainen, Jaramillo,
Näätänen, & Winkler, 1999).
Korzyukov, Winkler, Gumenyuk, Alho, and Näätä-
nen (2003) investigated whether this pitch-ascension
regularity can be reactivated similarly to regularities
based on primary sounds features. Seven tone-pairs
of ascending pitch varying randomly in their absolute
frequency levels were presented in the standard trains.
The retention interval was followed by a train starting
with one ascending-pitch (reminder) and a descend-
ing-pitch (deviant) tone pair. (Control sequences were
composed according to the scheme shown on the bot-
tom panel of Figure 1: pitch direction of the tone-
pairs varied randomly in the random-sound trains.) A
significant difference was observed between the re-
sponses elicited by the deviant and the corresponding
comparison tone-pair in the MMN latency range. The
scalp distribution of this potential difference matched
that of the MMN component elicited by descending-
pitch tone pairs presented infrequently amongst fre-
quent ascending-pitch tone pairs. This result demon-
strated that regularities based on secondary sound fea-
tures can be reactivated. Thus we can conclude that
the reactivation paradigm can indeed model real-life
situations in which recognition occurs on the basis of
acoustic subtleties.
2.3. Interpretation of the Reactivation
Phenomenon
Cowan et al. (1993) offered two alternative explana-
tions of their findings of “memory reactivation.” One
hypothesis suggests that during the retention interval,
the sensory memory traces involved in detecting
acoustic deviance enter a dormant state (i.e., a state in
which they cannot be directly accessed). The reminder
activates the corresponding dormant memory trace(s),
bringing them back to immediate memory (and thus
allowing the detection of the following deviants). The
alternative explanation assumes that, although the
memory traces required for deviance detection are
present and accessible, they are not consulted by the
MMN-generating process because the retention in-
terval causes a context change. That is, the sounds
presented after the relatively long retention interval
are not initially considered to be a continuation of the
preceding sound sequence, but rather the start of a
new sound group, whose regularities are yet to be de-
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
termined. In this explanation, the function of the re-
minder is to provide a link to the previous train,
reinstating its context as current and the related regu-
larities as relevant to the processing of the new
sounds.
The lack of MMN elicitation by the first sound of
the trains following the retention interval should thus
be taken either as a sign of the dormancy of the mem-
ory traces or as a sign that a change of context have
taken place. Based on the results of a behavioral test
and the analysis of the MMN studies known at the
time, Ritter and his colleagues (2002) argued that, in
just 10Ð15 seconds, auditory sensory memory traces
do not decay beyond usefulness. Evidence supporting
Ritter et al.’s conclusion has been obtained by Winkler
and his colleagues (2001; see also Gaeta et al., 2001),
who found that deviant tones may not elicit MMN at
the beginning of a short train when the trains are sepa-
rated by a silent interval of just 7 s duration. The trains
consisted of 4 tones of uniform stimulus duration (equi-
probably 100 or 300 ms) and were delivered equiproba-
bly with a stimulus onset asynchrony (SOA; onset-to-
onset interval) of 0.5 or 7 s. The silent interval sepa-
rating successive trains was always 7 s. The ERP re-
sponse to the first tone of those trains in which tone
duration differed from the preceding train was com-
pared with that elicited when tone duration matched
with the preceding train. The change in tone duration
elicited MMN in ca. half of the subjects when the
within-train SOA of the preceding train was 0.5 s. In
contrast, the same deviants following the same silent
interval (the between-train interval was always 7 s)
elicited MMN in all subjects when the within-train
SOA of the preceding train was 7 s (i.e., equal to the
inter-train interval). This result cannot be explained
on the basis of sensory memory alone, because, at the
time when the deviant tone was delivered, the sensory
memory trace of the standard tone must have been
stronger when the preceding train were delivered with
the short SOA than when it was delivered with the
long SOA. This is because more standard tones were
delivered with short than the long SOA within the last
ca. 10 s preceding the deviant tone. Therefore, we
must assume than the sensory memory trace of the
standard tone was present and accessible in both con-
ditions at the time when the deviant tone was deliv-
ered. Thus, this result demonstrates that the lack of
MMN cannot be taken as proving that no sensory
memory trace can be accessed by the MMN-generat-
ing process. Winkler et al. (and also Gaeta et al.) inter-
preted their result in terms of the context change hy-
pothesis. On the basis of these results, Ritter et al.
(2002) favored the context reactivation (or reinstate-
ment) explanation of the MMN reactivation phenome-
non.

10 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
However, since then, Winkler et al. (2002) showed
reactivation after a much longer (30 s) retention in-
terval and that reactivation occurred also in those sub-
jects who could not discriminate the standard and de-
viant tones when these were separated by 30 s of si-
lence. The latter result is all the more important since
Keller et al. (1995) found a small effect of tone re-
hearsal on tone comparisons performance for over-10-
s retention intervals. This may explain why Ritter et
al. (2002) as well as our own test (see the next section)
found significant residual memory after 11Ð15 s in
active tone comparison tasks. However, the finding of
reactivation in subjects who could not perform the
comparable discrimination task suggests that reactiva-
tion may involve both of the processes brought up by
Cowan et al. (1993): The reminder connects the new
sounds to a previous context (reinstatement) as well
as reactivating the possibly dormant memory repre-
sentations that describe the regular characteristics of
this context.
2.4. Reactivation in an Active
Tone-Comparison Experiment
The comparisons between results obtained with MMN
and in behavioral studies (discussed in the previous
section) make it imperative to test reactivation in an
active paradigm that is analogous to the ones tested in
the passive situation. Although the reactivation para-
digm was intended to model situations in which re-
cognition occurs without voluntary effort, one would
assume that reactivation should also occur in situa-
tions in which the subject actively tries to maintain
some sensory information. This was tested in two ex-
periments in which subjects were required to judge
whether a test tone was the same or different com-
pared with a previously presented standard tone. The
two experiments differed only in whether the standard
tone was presented only once (1-Standard Experi-
ment; 21 participants, 9 male, 16Ð23 years of age,
19.4 years mean age) or six times in a row with 0.75 s
SOA (6-Standard Experiment; 25 participants, 13
male, 18Ð31 years of age, 22.4 years mean age).
2.4.1. Stimuli and Procedure
Eighteen sets of tones and a burst of white noise, band-
pass filtered between 300 and 1500 Hz, were generated.
The duration of all sounds was 250ms (including 5ms
rise and 5ms fall times), their intensity 70 dB (SPL).
Each tone set consisted of 3 tones separated in fre-
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
quency by proportionally equal steps of 2.5, 3, 3.5, 4,
or 5 % (see later). (For later reference, the middle tone
of the sets will be denoted as “A” as it was always the
first to be presented; the lower-pitched one will be
denoted as “B,” the higher as “C,” and the aperiodic
noise as “N”). The frequency of the middle (A) tone
of the lowest set was 400 Hz; the middle tones of
neighboring sets were separated by 6 % in frequency.
Each trial presented tones from only one set. Tone sets
appeared in the trials in a randomized order and with
equal probability. Trials started with the standard
tone(s), which was always the middle tone of a tone
set (“A”), followed by a silent retention interval,
which was, with equal probability, 11, 12, 13, 14, or
15 s. The sound following the retention interval was
selected with equal (25 %) probability from the three
tones of the set and the noise-burst. These second
tones were considered to be reminders (A valid; B, C,
and N invalid). Following the reminder by an SOA of
0.75 s, the test tone was delivered. The test tone was
equiprobably chosen from the three tones (A, B, and
C) of the set selected for the trial. Thus, 12 types of
trials were presented (the order of the sounds being
standard, reminder, test): AAA, AAB, AAC, ABA,
ABB, ABC, ACA, ACB, ACC, ANA, ANB, and
ANC. Each trial type occurred 18 times, each time
based on a different tone set (216 trials, overall). Sub-
jects were instructed to press a button if they thought
that the test tone was identical to the standard tone(s)
and a different button if they thought that the two
tones were different. The reminder (the sound preced-
ing the test tone) was introduced to them as a warning
signal preparing them for the delivery of the test tone.
They were informed that the warning sound carried
no information about the task and so they should not
rely on it in their judgment. The instructions empha-
sized the requirement of correct responses and placed
no time pressure on giving the response. Subjects
were also motivated by a scheme of performance-de-
pendent bonus payments to do the task as best they
could. Subjects started each trial when they felt ready
for it.
Prior to starting the main test, we established the
frequency difference at which subjects could reliably
(⬎ 90 %) discriminate two tones of slightly different
frequencies. Subjects were presented with pairs of
tones separated by a 2-s SOA. The two tones of the
pair were either identical or slightly different from
each other in frequency. In separate stimulus blocks
(10 pairs per block), the frequency difference was
either 2.5, 3.0, 3.5, 4.0, or 5.0 %. Absolute tone fre-
quencies varied across the 18 preselected frequency
sets (see above). Testing started with the highest (5 %)
frequency separation. After each successful stimulus

11I. Winkler & N. Cowan: From Sensory to Long-Term Memory
block (a block in which at least 9 of the 10 responses
were correct), frequency separation was decreased.
Frequency separation was increased after two succes-
sive blocks in which performance did not reach the
criterion level. Testing was finished when a frequency
separation was found at which the subject performed
at the desired level, but had to turn back twice from
the one-step lower frequency-separation level. On
average, subjects of the 1-standard experiment needed
4.1 %, whereas subjects of the 6-standards experiment
needed 3.8 % frequency separation for reliable dis-
crimination performance. Frequency separation be-
tween the B and A and A and C tones was set up in
the main experiment to equal the level established in
the preliminary testing. Subjects then received train-
ing in the task of the main experiment. Two blocks of
18 trials each were presented to them with the reten-
tion interval set to 3.5 s and feedback given after each
response. All other parameters were identical to the
corresponding ones in the main experiment.
2.4.2. Model for Analyzing the Results
Performance was analyzed with the help of a mathe-
matical model
2
of performance that assumed additiv-
ity between the following effects:
(1) Memory (“M”) Ð Performance based on the sen-
sory memory trace present at the time the warning
(reminder) tone was presented.
(2) Reactivation (“R”) Ð Only present on AAX type
of trials (X can be either A, B, or C); increases
performance.
(3) Interference from the warning tone (“I”) Ð
Decreases performance by degrading the residual
memory trace. Interference was assumed to be
zero for noise (ANX) trials. This assumption may
not be accurate. As a consequence, “M” will be
slightly underestimated.
(4) Strategy to answer according to the relationship
between the warning and the test tone (“S”) Ð
Using this strategy boosts performance on some
types of trials, such as ABC, but degrades perfor-
2
The formal model of performance is as follows. P(AXY) stands for the performance in the AXY trials (X can be A, B, C, or
N and Y can be A, B, or C); for other abbreviations, see the text. Equations are numbered from (1) to (7).
(1) P(ANA)=M+Bn
(2) [P(ANB) + P(ANC)]/2 = M - Bn
(3) P(AAA)=M+R+S+Ba- I
(4) [P(AAB) + P(AAC)]/2=M+R+S- Ba - I
(5) [P(ABA) + P(ACA)]/2 = M - S + Bbc - I
(6) [P(ABB) + P(ACC)]/2 = M - S - Bbc - I
(7) [P(ABC) + P(ACB)]/2=M+S- Bbc - I
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
mance on others, such as ABB. The frequency
separation between the warning and the test tones
was larger in the ABC and ACB trials than in any
of the other trials. This may have increased the
likelihood in these types of trials that answers were
given on the basis of the relationship of the warn-
ing and the test tones. By assuming that “S” was
equal in all trial types, we may thus underestimate
“S” for the ABC and ACB type of trials and, as a
consequence, underestimate the size of the reacti-
vation (“R”) effect.
(5) Bias to answer “equal” over answering “different”
(“B”) Ð Bias boosts performance on some types
of trials, such as ABA, but degrades performance
on others, such as ABB. This tendency was as-
sumed to depend on the relationship between the
standard tone and the warning sound. Therefore
three separate “B” values were considered, one for
the AAX trials, another for the ABX and ACX
trials, and the third for the ANX trials: Bias vari-
ables were named Ba, Bbc, and Bn, respectively.
The number of variables (effects) in the model
equaled the number of independent measurements.
Therefore, the effect values (the amount by which
each effect contributed to performance) could be un-
ambiguously determined, separately for the 1- and
6-Standard experiments.
2.4.3. Results and Discussion
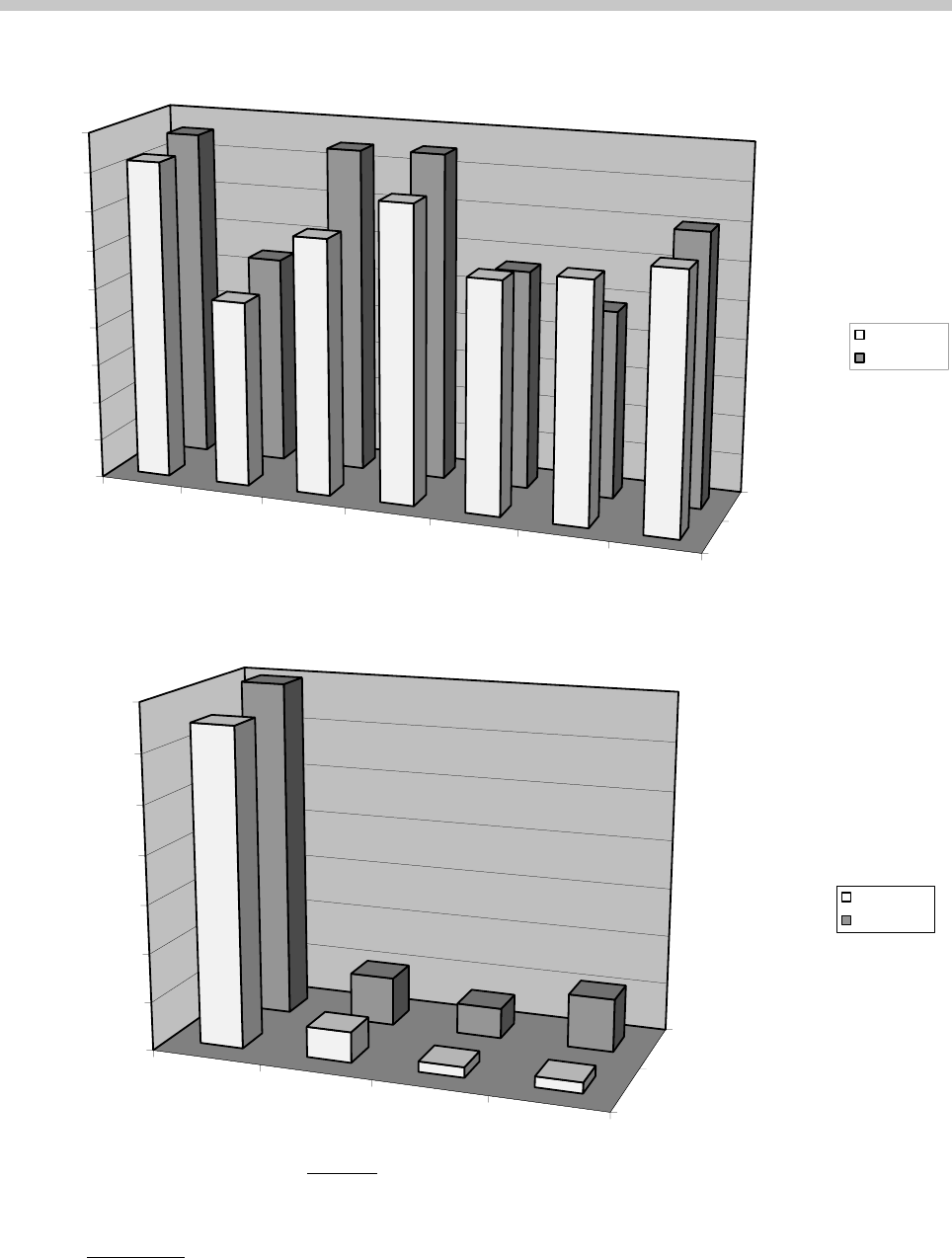
Figure 3 presents the results of both active reactivation
experiments (hit percentages according to trial cate-
gories on the top panel and the calculated model val-
ues on the bottom panel). Although the memory factor
was responsible for the largest segment of perfor-
mance, reactivation proved to be significant in both
experiments (p ⬍ 0.05 in the 1-Standard Experiment
and p ⬍ 0.01 in the 6-Standard Experiment; one-
group Student’s t tests). The strategy and interference
factors were found to be significant only in the 6-
Standard Experiment (p ⬍ 0.01 and p ⬍ 0.05, respec-
tively). No significant differences were found between
the two experiments for any of the model factors (dif-
12 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
ANA
ANY
AAA
AAY
AYA
AYY
AYZ
1-Standard
6-Standard
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
Hit %
Trial types
Tone Comparison
1-Standard
6-Standard
Memory
Reactivation
Interference
Strategy
1-Standard
6-Standard
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
Hit %
Model Values
1-Standard
6-Standard
Figure 3. Active reactivation experiments. Top panel: The percentage of correct responses (hits), separately for the 1-Standard (front
row) and 6-Standard (back row) experiments and sorted by trial categories. The 12 trial types (see the text) were analyzed in 7
categories created according to the different effects included in the model. ANY presents the average performance in the ANB and
ANC trials, AAY the AAB and AAC trials, AYA the ABA and ACA trials, AYY the ABB and ACC trials, and AYZ the ABC and
ACB trials. Bottom panel: Contribution of the modeled effects (in hit percentage) to the performance in the two experiments. Note
that the interference factor decreases performance (all other factors, when their value is positive, increase performance).
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers

13I. Winkler & N. Cowan: From Sensory to Long-Term Memory
ference in the strategy factor was marginally signifi-
cant: F(1, 44) = 3.80, p = 0.0575; one-way between-
group ANOVA). In the 6-Standard but not in the 1-
Standard Experiment, reactivation was positively
correlated with memory (Spearman rank correlation
0.44, p ⬍ 0.05), whereas strategy was negatively
correlated with memory (-0.43, p ⬍ 0.05). These re-
sults suggest that reactivation occurs also when sub-
jects actively maintain auditory sensory information,
although its effect is relatively small as long as the
residual memory is still sufficiently strong (i.e., in the
present situation, over 60 % of the performance can
be attributed to residual memory, whereas only ca.
10 % to reactivation).
3. General Discussion:
A Reevaluation of Sensory Memory
3.1. What Kind of Memory Representations
Are Reactivated?
MMN elicitation is based on a memory representation
describing auditory regularities, not just auditory sen-
sory memory traces. This has been shown by the re-
sults of those studies that tested violations of various
nonrepetitive regularities (for a review, see Näätänen
et al., 2001). However, these memory representations
are not independent of the concrete sound feature
levels even in the most “abstract” cases. For example,
Paavilainen, Simola, Jaramillo, Näätänen, and Win-
kler (2001) presented subjects with tones varying in
frequency and intensity. Most tones conformed to a
rule, which was, in separate stimulus blocks, either
“the higher the frequency the higher the intensity” or
“the higher the frequency the lower the intensity”. In-
frequent tones violating the standard rule (high-fre-
quency soft and low-frequency loud tones or high-fre-
quency loud and low-frequency soft tones, depending
on the rule) elicited MMN. This result suggests that
the memory representation involved in MMN genera-
tion encoded the abstract feature-conjunction regular-
ity. One aspect of the results, however, suggested that
the representation of the regularity was not fully inde-
pendent of the actual levels of the relevant auditory
features. The MMN amplitude was somewhat higher
in response to deviants that fell farther from the center
of the feature distribution of the regular sounds.
(However, no regular tone elicited the MMN, not even
the ones with extreme levels in both features.) The
MMN-amplitude differences suggest that the standard
tones of medium frequency and medium intensity
were regarded as a prototype within the abstract regu-
larity.
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
The picture emerging from the MMN literature is
that the representations of the stimulation must in-
clude not only regularities, but also certain feature val-
ues that serve as reference points or anchors. An an-
chor point would allow an abstract rule or regularity
to be translated into specific feature values. That way,
the same regularity could be associated with different
anchors in different stimulus situations, allowing for
the efficient storage of information and the possibility
to maintain alternative descriptions of the same se-
quence of sounds. Simultaneous representation of
multiple redundant regularities describing the same
sound sequence has been demonstrated for MMN gen-
eration (Horva
´
th et al., 2001). Horva
´
th and his col-
leagues presented a regular sequence of tones alternat-
ing in pitch (ABABAB . . ., where A and B are two
tones differing only in pitch). The presence of repre-
sentations for different redundant regularities was
tested by presenting deviants that violated one pos-
sible description (rule) while conforming to a different
description of pitch alternation. It was found that
memory representations of at least one “local” rule (A
tone is always followed by B and vice verse) and one
“global” rule (every second tone is A and every other
is B) are simultaneously maintained and incoming
sounds are checked against them in parallel. Further
results suggested that possibly also representations of
more general versions of alternation are kept active at
the same time (rules, such as Higher toneÐLower
toneÐHigher toneÐLower tone . . . and alternation
with variable interstimulus intervals). If, as was shown
by Horva
´
th et al. (2001), the auditory system main-
tains multiple redundant representations even for sim-
ple acoustic regularities, concise abstract regularity
descriptions actualized with a minimal amount of sen-
sory data offer an economical form of information
storage.
The hypothesized structure of regularity represen-
tations suggests that we can reactivate sensory infor-
mation that was encountered within a context with dis-
tinctive regular characteristics. A stimulus that con-
forms these regularities serves as a reminder, activa-
ting the regularity representations and thus bringing
the corresponding context to immediate memory. Re-
activation of the characteristic features of an object
could serve as the basis of its recognition. Features of
the reactivation process are in perfect correspondence
with everyday experience of object recognition. We
can easily recognize objects appearing in their usual
context. The more experience we have with the given
object and the more distinctive its sensory features the
more likely that we can recognize it.
The characteristics of the memory representations
as shown by the reviewed reactivation experiments,

14 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
i.e., resistance to decay and interference, encoding of
higher-order regularities extracted from acoustical
variance, are compatible with the assumed characteris-
tics of long-term memory records. Thus it seems pos-
sible that in the auditory modality, the regularity rep-
resentations indexed by the MMN component form
the link between immediate sensory memory and
long-term storage of sensory information.
3.2. What Distinguishes Auditory Sensory
Memory from Categorical Memory?
The MMN and new behavioral results discussed above
help to establish several points about memory for
acoustic stimuli. First, the regularities of the acoustic
pattern, including grouping and organization of acous-
tic information, are important for memory. Second,
aspects of these regularities can be stored in long-term
memory and reactivated later. Given these results, sev-
eral issues arise with respect to behavioral research on
memory and the models that have been based on it.
First, does it still make sense to think of this informa-
tion as especially “sensory,” as opposed to categorical,
in nature? Second, what types of overall models of
processing can represent all of the results?
In the following we shall re-examine the evidence
supporting the distinction of sensory memory traces
form categorical memory storage for each of the four
critical qualities mentioned in the introduction:
(1) sensory memory does not depend on attention,
(2) it is modality-specific, (3) it includes information
that is finer than any meaningful set of categories, and
(4) it is short-lived. How do these qualities apply to
the retention of acoustic regularities?
3.2.1. Attention-Independence
A traditional objection to the attention-independence
of sensory processing comes from findings showing
attentional modulation of neural signals associated
with early afferent processing. Results showing that
mid-latency ERP components, such as the auditory
P50 wave are attenuated when attention is directed
away from the stimuli have been interpreted in terms
of “sensory gating” (e.g., Guterman, Josiassen, & Ba-
shore, 1992). It has been suggested that the afferent
flow of sensory information is under control from the
prefrontal cortex, a structure linked with the voluntary
direction of behavior (Knight, Staines, Swick, &
Chao, 1999). The notion of sensory gating may be
compatible with the attentional spotlight theory. Sup-
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
porting results for the auditory modality have been
obtained in experiments investigating the detection of
auditory targets as a function of the target’s distance
from a precued (expected) location. A graded
decrease of accuracy and slower reaction times have
been found with increased distance between the target
and the precued location (Arbogast & Kidd, 2000;
Mondor & Zatorre, 1995; Rorden & Driver, 2001).
Comparable amplitude gradients have been found
both for some of the early, exogenous (N1) as well
as for later, endogenous (MMN, P3) ERP responses
(Arnott & Alain, 2002; Teder-Salejärvi, Hillyard,
Roder, & Neville, 1999).
Effects in sound organization provide another line
of argument against the attention-independence of au-
ditory sensory memory. Bregman (1990) suggested
that several heuristic processes analyze the auditory
input in parallel. These processes provide alternative
solutions to breaking down the complex acoustic input
into coherent sequences of sound. When two strong
alternatives of sound organization emerge from the
initial analysis (ambiguous auditory scenes), one can
voluntarily choose between them. Since the auditory
information stored about sounds depend on how they
are organized (Dowling, 1973), ambiguous auditory
scenes allow voluntary modulation of auditory sen-
sory memory. In a similar vein, Sussman et al. (2002)
found that informing subjects about the large-scale
structure of a sound sequence determined what sounds
are detected as deviants in a sound sequence. Sussman
et al. presented a tone sequence composed of a repeat-
ing tone pattern (AAAABAAAAB . . .; where B was
higher in pitch than A). Tones were presented at an
intermediate (700ms) SOA, because previous studies
showed that at shorter SOAs (100ms), the sequence is
unambiguously represented as a repeating tone pattern
(Sussman, Ritter, & Vaughan, 1998), whereas at
longer SOAs the large-scale regularity is not automati-
cally detected (Scherg, Vajsar, & Picton, 1989). Of the
A tones, 2.5 % were exchanged for a lower-pitched (C)
tone. When subjects were instructed to press a re-
sponse button for the rare low-pitched C tones MMN
was elicited by the B tones, which were relatively rare
(appeared in 20 % of the time) compared with the A
tones. In contrast, when subjects were informed about
the regularly repeating tone pattern and were in-
structed to press the response button whenever the
pattern was broken (that is, to the same C tones), the
B tones did not elicit MMN. The lack of MMN by the
B tones indicates that the tone sequence was repre-
sented in terms of the repeating tone pattern. In this
case, the B tone is part of the repeating pattern and,
therefore, it does not violate a regularity. Thus it ap-
pears that, at least in certain cases, auditory memory

15I. Winkler & N. Cowan: From Sensory to Long-Term Memory
can be influenced by top-down processes. Moreover,
some authors suggest that even the unambiguous cases
of sound organization require attention (e.g., Carlyon,
Cusack, Foxton, & Robertson, 2001), although this is-
sue is still controversial (see, e.g., Winkler et al.,
2003).
Taken together, this literature does call into ques-
tion whether complex acoustic patterns are held in a
way that is independent of attention; but it also calls
into question the more fundamental assumption that
sensory memory is held in a manner that is independ-
ent of attention. That never has been rigorously exam-
ined, and the aforementioned study by Keller et al.
(1995) did find that attention had an effect on memory
for tone frequencies necessary for comparisons within
a musical category.
3.2.2. Modality-Specificity
Regarding the second criterion of sensory memory, its
modality-specificity, here the results seem unequivo-
cal. Frankish (1985) and Cowan, Saults, Elliott, and
Moreno (2002) compared memory for grouped and
ungrouped lists of spoken or printed verbal items. The
result in both studies was that grouping has a much
larger, beneficial effect in the auditory modality. In
fact, Cowan et al. found that, with visual pre-
sentations, grouping could be accomplished just as
well on trials in which the stimuli themselves were
presented at a steady pace as on trials in which the
stimuli were temporally grouped into 3 clearly distinct
sets of 3 digits. Both presentation schedules produced
serial recall results for the 9-digit lists that were
slightly scalloped in a manner, suggesting that the 9
digits had been mentally grouped accordingly. (The
stimulus grouping on some trials apparently induced
a similar mental grouping on the remaining trials with
visual presentation.) In contrast, with auditory pre-
sentation of digits, the outcome for grouped lists was
far superior to the outcome for lists of digits presented
at a steady pace. It is as if the acoustic modality, un-
like vision, inherently carries timing information that
cannot be ignored within the mental representation.
In accordance with this notion, Näätänen & Winkler
(1999) suggested that the time-line of acoustic stimu-
lation serves as the core for integrating the outcome
of the various auditory feature analyzers into a unitary
representation of the auditory stimulus: “We suggest
that the critical step between the stage of fragmentary
stimulus information (maintained in the feature traces)
and the emergence of the auditory stimulus represen-
tation is the synthesis of the static stimulus features
with the temporal envelope of the stimulus event. An
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
auditory stimulus cannot be fully described by static
features alone. Therefore, in the formation of an audi-
tory stimulus representation, the extracted features
must be aligned with the passage of time.” (Näätä-
nen & Winkler, 1999, p. 848).
3.2.3. Resolution
The third traditional hallmark of sensory information
is that it is more fine-grained than categorical infor-
mation. The question is whether this fine-grained in-
formation is present within the same memory repre-
sentation that stores extended acoustic patterns. Evi-
dence to that effect is that the ability to detect a differ-
ence between two tones differing less than a musical
note category is affected by the pattern of prior tone
presentations. Cowan, Saults, and Nugent (1997) var-
ied not only the time between tones to be compared,
but also the time between trials. It was found that
when trials were further apart, performance improved.
One way to explain that result is that, when trials are
too close together, the first tone in the current pair
may be grouped in memory with tones from the pre-
ceding trial. So, it appears that there is no such thing
as a simple, pure acoustic memory for a single tone
separate from the pattern of tones that has formed.
3.2.4. Duration
The fourth typical characteristic of sensory memory
was its short life. However, we already have discussed
evidence that there is memory for acoustic properties
that lasts a long time, such as information about a
friend’s voice. There is abundant evidence for long-
term auditory memory that allows a recall advantage
over visual presentation, even after an intervening dis-
tracting task, provided that the items to be recalled
also are separated by other items. Research on long-
term modality effects recently was reviewed by Gardi-
ner and Cowan (2003). Also, Cowan, Saults, and Nu-
gent (2001) reanalyzed the evidence on tone compari-
sons by Cowan et al. (1997) and found that, under
certain circumstances in which the separation between
trials was the largest, it was unclear if there was any
loss of information at all as a function of the time
between tones to be compared. In other words, the
“decay” of auditory sensory information across sev-
eral seconds may actually be a matter of the tone be-
coming more and more confusable with tones from
previous trials, as opposed to an actual loss of sensory
persistence as has typically been assumed. However,
this dramatic conclusion awaits further testing.

16 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Although these findings suggest that sensory infor-
mation lasts in memory rather than decaying rapidly,
there are some apparent contradictory findings. Some
studies have shown that the final serial position of
acoustically presented lists carries information that is
especially vivid and may be represented differently
from items presented in any other position of the list.
There has been some debate on this point (cf. Ba-
lota & Engle, 1981; Bloom & Watkins, 1999). How-
ever, Cowan, Nugent, Elliott, and Saults (2000) found
a clear distinction of the final serial position in a de-
velopmental study of memory for digit lists that were
ignored during their presentation while a silent game
involving rhyming pictures was played. Occasionally,
the rhyming game was interrupted and the computer
keyboard was to be used to recall the most recent spo-
ken list. The delay between the end of this last list and
the recall cue was 1, 5, or 10 s. For most serial posi-
tions, children of two ages and adults all forgot the
items at the same rate across the variable retention
interval. For the final serial position, though, the result
was strikingly different than for any other serial posi-
tion. Young children (in second grade) forgot this final
item much more rapidly than subjects in the older
groups.
There is another finding suggesting that there may
be a special form or function of memory that is spe-
cific to the final item (although see Bloom & Watkins,
1999 for evidence that it is not quite that specific). It
has been obtained in studies of the suffix effect, in
which a final list item that is not to be recalled (e.g.,
the word “go”) interferes with memory for items at
the end of the list (Crowder & Morton, 1969; Morton,
Crowder, & Prussin, 1971). Balota and Ducek (1986)
presented a suffix immediately after a spoken list or
after a 20-second delay and found that there was a
suffix effect in either case, but that the similarity be-
tween the voice of the speaker of the list and suffix
mattered only if the suffix was presented immediately.
This again suggests that there is some specific acous-
tic information that is very fragile and some broader
pattern of information that is more durable. It is un-
clear if the specific acoustic information disappears
as a function of absolute time or, more in keeping
with other evidence we have presented, disappears as
a function of the shifting context; the specific acoustic
information apparently would lose its contextual rele-
vance especially quickly over time.
3.3. The “Regularity-Record Plus Anchor”
Hypothesis
The theoretical need, then, is to tie together findings
suggesting that acoustic information is retained for a
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
long time and the findings suggesting that the final
item may be of special importance. Information on
long-term retention includes, for example, the mis-
match negativity results showing that patterns are
taken into account and that rapid decay does not de-
scribe performance well (e.g., Winkler et al., 2001),
and behavioral results on long-term modality effects
(e.g., Gardiner & Cowan, 2003) and sensory memory
stability (Cowan et al., 2001). Information on the spe-
cial importance of the most recent item includes the
role of this item as an anchor, or as a reminding or
reactivating stimulus, in mismatch negativity pro-
cedures (Cowan et al., 1993; Korzyukov et al., 2003;
Winkler et al., 1996) and, behaviorally, the develop-
mental difference in memory decay for the list-final
item only (Cowan et al., 2000) and the tendency of
modality and suffix effects to be largest at the end of
the list.
One way to combine these factors theoretically is
with the notion that acoustic memory is a record of
regularities, and that the last item serves as an anchor
for this record. As an analogy, every member of an
orchestra has a mental record of the music that is to
be played, but still needs a common pitch to anchor
the memory and tune the instrument before the musi-
cal piece is played. To carry the analogy further, the
melody can be held in long-term memory but the an-
choring note must be played on the spot for it to be
of any use (except for musicians with the absolute
pitch ability). If an intruder came into the room during
tuning and played a deviant note, the process would
be corrupted and the correct anchor probably would
have to be repeated. That is analogous to the use of a
reactivating reminder in the mismatch negativity pro-
cedure and to the corrupting effect of a suffix in list-
recall procedures.
Characteristics of the suffix effect are compatible
with the above-suggested structure of auditory mem-
ory (i.e., regularity plus anchor). Most studies of the
suffix effect show that it can occur even when the
suffix remains the same from trial to trial. However,
the situation is different when list items are separated
by distracting tasks, making them temporally very dis-
tinct and producing long-term modality effects. Under
that situation, a suffix effect eliminating the auditory
modality advantage occurs, but only if a different suf-
fix is used on every trial (Glenberg, 1984). In the
short term, the most recently presented acoustic item
may have a special vividness that is susceptible to in-
terference from any other sound. In the long term,
though, the most recent item may carry acoustic infor-
mation that reminds the subject of the rest of the
acoustically presented list. When that item was a suf-
fix that was not to be recalled, it may detract from the

17I. Winkler & N. Cowan: From Sensory to Long-Term Memory
power of the list-final item to serve as a unique anchor
for the list.
3.4. Considering some Current
Memory Models
Last, we can ask the question of what established
models can or cannot handle these results. The fore-
most model of temporary memory, currently, is the
working-memory model of Baddeley (1986). That
model postulates the existence of two buffer stores, a
phonological buffer and a visuospatial buffer. Re-
cently, a short-term episodic buffer also has been pos-
tulated (Baddeley, 2000). However, in our view, none
of these buffers make sufficient accommodation for
specifically sensory information. The phonological
buffer cannot be used to explain the auditory modality
superiority effect. To explain it, it seems necessary to
postulate the existence of sensory memory stores in
addition to the more abstract, phonological, and spa-
tial representations that are highlighted in Baddeley’s
model.
In the similar but alternative model of Cowan
(1988, 1995, 1999), sensory memory and categorical
memory both are considered to reflect activated sub-
sets of the long-term memory system (though new
links are formed between items that are attended con-
currently, and these new links then become part of the
long-term memory system). Results showing fast
task-independent effects of information stored in
long-term memory records on auditory change detec-
tion are fully compatible with Cowan’s model. For ex-
ample, phonetic category boundaries and prototypes
have been shown to affect the MMN response elicited
in ignored sequences of speech stimuli (Aaltonen,
Eerola, Hellstrom, Usipaikka, & Lang, 1997;
Näätänen et al., 1997; Winkler et al., 1999). In Cow-
an’s model, the modality-specific effects are handled
by assuming that there will be interference between
the activated representation of a recently presented
item and a new stimulus when the two share similar
features. Cowan’s (1988, 1995, 1999) model does not
specifically explain why the auditory modality superi-
ority effect occurs; nor does it specify the nature of
memory for regularities in the auditory memory sys-
tem. Yet, it seems more open to the possibility of sepa-
rate acoustic and phonological information sources
than does Baddeley’s (1986, 2000) model.
Given the way in which the expectations of the
earlier models of processing have failed, it is perhaps
not surprising to learn that some researchers have pro-
moted models in which there is no distinction at all
between shorter-term and longer-term memory (e.g.,
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
Nairne, 2002). Such models are well suited to accom-
modate the finding that memory for acoustic regulari-
ties is long lasting, and to accommodate similarities
between the results of ostensibly short-term and long-
term memory phenomena. However, such models do
not yet appear to have a ready explanation for differ-
ences that are observed between shorter-term and
longer-term memory phenomena. One issue here is
what we mean by the term “reactivation.” It carries
with it the assumption that there is such a thing as
activation. If there is no distinct form of memory in
the short term, then there is no such thing as tempo-
rary memory activation. Instead, what we term “acti-
vated” would actually be “relevant to the present
context.” In that case, reactivation effects would have
to be more accurately portrayed as reminder effects.
Ultimately, new modeling efforts will be needed to
explain precisely how it is that regularities can be
stored in acoustic memory, and to determine whether
some minimal amount of attention is needed to assist
in the formation of the memory for regularities that
is stored and later can be reactivated by a reminder
stimulus.
Acknowledgement
Due to space constraints this article has been pub-
lished outside of the special issue “Working Memory
and Cognition” (Issue 4, 2004) for which it was origi-
nally accepted.
This research was supported by the Hungarian Na-
tional Research fund grant OTKA T034112 and U.S.
National Institutes of Health grant HD-21338. We
thank Lı
´
via Pato
´
for conducting the behavioral reacti-
vation experiments.
References
Aaltonen, O., Eerola, O., Hellstrom, A
˚
., Uusipaikka, E., &
Lang, A. H. (1997). Perceptual magnet effect in the light
of behavioral and psychophysiological data. Journal of the
Acoustical Society of America, 101, 1090Ð1106.
Arbogast, T. L., & Kidd, G. (2000). Evidence for spatial tuning
in informational masking using the probe-signal method.
Journal of the Acoustical Society of America, 108, 1803Ð
1810.
Arnott, S. R., & Alain, C. (2002). Stepping out of the spotlight:
MMN attenuation as a function of distance from the attended
location. NeuroReport, 13, 2209Ð2212.
Atkinson, R. C., & Shiffrin, R. M. (1968). Human memory: A
proposed system and its control processes. In K. W.
Spence & J. T. Spence (Eds.), The psychology of learning
and motivation: Advances in research and theory (Vol. 2,
pp. 89Ð195). New York: Academic Press.

18 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Baddeley, A. D. (1986). Working memory. Oxford Psychology
Series #11. Oxford: Clarendon Press.
Baddeley, A. (2000). The episodic buffer: a new component of
working memory? Trends in Cognitive Sciences, 4, 417Ð
423.
Balota, D. A., & Duchek, J. M. (1986). Voice-specific informa-
tion and the 20-second delayed suffix effect. Journal of Ex-
perimental Psychology: Learning, Memory, and Cognition,
12, 509Ð516.
Balota, D. A., & Engle, R. W. (1981). Structural and strategic
factors in the stimulus suffix effect. Journal of Verbal Learn-
ing and Verbal Behavior, 20, 346Ð357.
Bloom, L. C., & Watkins, M. J. (1999). Two-component theory
of the suffix effect: Contrary findings. Journal of Experi-
mental Psychology: Learning, Memory, and Cognition, 25,
1452Ð1475.
Bregman, A. S. (1990). Auditory Scene Analysis: The Percep-
tual Organization of Sound. Cambridge, MA: MIT Press.
Broadbent, D. E. (1958). Perception and communication.New
York: Pergamon Press.
Carlyon, R. P., Cusack, R., Foxton, J. M., & Robertson, I. J.
(2001). Effects of attention and unilateral neglect on audi-
tory stream segregation. Journal of Experimental Psychol-
ogy: Human Perception and Performance, 27, 115Ð127.
Cowan, N. (1984). On short and long auditory stores. Psycho-
logical Bulletin, 96, 341Ð370.
Cowan, N. (1988). Evolving conceptions of memory storage,
selective attention, and their mutual constraints within the
human information-processing system. Psychological Bulle-
tin, 104, 163Ð191.
Cowan, N. (1995). Attention and memory. An integrated frame-
work. Oxford: Oxford University Press.
Cowan, N. (1999). An embedded-processes model of working
memory. In A. Miyake & P. Shah (eds.), Models of Working
Memory: Mechanisms of active maintenance and executive
control (pp. 62Ð101). Cambridge, UK: Cambridge Univer-
sity Press.
Cowan, N., Nugent, L. D., Elliott, E. M., & Saults, J. S. (2000).
Persistence of memory for ignored lists of digits: Areas of
developmental constancy and change. Journal of Experimen-
tal Child Psychology, 76, 151Ð172.
Cowan, N., Saults, J. S., Elliott, E. M., & Moreno, M. (2002).
Deconfounding serial recall. Journal of Memory and Lan-
guage, 46, 153Ð177.
Cowan, N., Saults, J. S., & Nugent, L. D. (1997). The role of
absolute and relative amounts of time in forgetting within
immediate memory: The case of tone pitch comparisons.
Psychonomic Bulletin & Review, 4, 393Ð397.
Cowan, N., Saults, S., & Nugent, L. (2001). The ravages of
absolute and relative amounts of time on memory. In H. L.
Roediger III, J. S. Nairne, I. Neath, & A. Surprenant (eds.),
The nature of remembering: Essays in honor of Robert G.
Crowder (pp. 315Ð330). Washington, DC: American Psy-
chological Association.
Cowan, N., Winkler, I., Teder, W., & Näätänen, R. (1993).
Short- and long-term prerequisites of the mismatch negativ-
ity in the auditory event-related potential (ERP). Journal of
Experimental Psychology: Learning, Memory, and Cogni-
tion, 19, 909Ð
921.
Craik, F. I. M., & Kirsner, K. (1974). The effect of speaker’s
voice on word recognition. Quarterly Journal of Experimen-
tal Psychology, 26, 274Ð284.
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
Crowder, R. G. (1989). Imagery for musical timbre. Journal of
Experimental Psychology: Human Perception and Perfor-
mance, 15, 472Ð478.
Crowder, R. G., & Morton, J. (1969). Precategorical acoustic
storage. Perception & Psychophysics, 5, 365Ð373.
Deutsch, D. (1975). The organization of short-term memory for
a single acoustic attribute. In D. Deutsch & J. A. Deutsch
(eds.), Short-term memory (pp. 107Ð151). New York: Aca-
demic Press.
Dowling, W. J. (1973). Rhythmic groups and subjective chunks
in memory for melodies. Perception & Psychophysics, 14,
37Ð40.
Durlach, N. I., & Braida, L. D. (1969). Intensity perception:
I. Preliminary theory of intensity resolution. Journal of the
Acoustical Society of America, 46, 372Ð383.
Frankish, C. (1985). Modality-specific grouping effects in
short-term memory. Journal of Memory and Language, 24,
200Ð209.
Fujisaki, H., & Kawashima, T. (1971). A model of the mecha-
nisms for speech perception - quantitative analysis of cate-
gorical effects in discrimination. Annual Report of the Engi-
neering Research Institute of the Faculty of Engineering,
University of Tokyo, 30, 59Ð68.
Gaeta, H., Friedman, D., Ritter, W., & Cheng, J. (2001). The
effect of perceptual grouping on the mismatch negativity.
Psychophysiology, 38, 316Ð324.
Gardiner, J. M., & Cowan, N. (2003). Modality effects. In J. H.
Byrne, H. Eichenbaum, H. Roediger III, & R. F. Thompson
(eds.), Learning and Memory (2nd edition) (pp. 397Ð400).
New York: Macmillan.
Glenberg, A. M. (1984). A retrieval account of the long-term
modality effect. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 10, 16Ð31.
Guterman, Y., Josiassen, R. C., & Bashore, T. R. Jr (1992).
Attentional influence on the P50 component of the auditory
event-related brain potential. International Journal of Psy-
chophysiology, 12, 197Ð209.
Halgren, E., Baudena, P., Clarke, J. M., Heit, G., Liegeois, C.,
Chauvel, P., & Musolino, A. (1995). Intracerebral potentials
to rare target and distractor auditory and visual stimuli. I.
Superior temporal plane and parietal lobe. Electroencepha-
lography & Clinical Neurophysiology, 94, 191Ð220.
Horva
´
th, J., Czigler, I., Sussman, E., & Winkler, I. (2001).
Simultaneously active pre-attentive representations of local
and global rules for sound sequences. Cognitive Brain Re-
search, 12, 131Ð144.
Keller, T. A., Cowan, N., & Saults, J. S. (1995). Can auditory
memory for tone pitch be rehearsed? Journal of Experimen-
tal Psychology: Learning, Memory, and Cognition, 21, 635Ð
645.
Knight, R. T., Staines, W. R., Swick, D., & Chao, L. L. (1999).
Prefrontal cortex regulates inhibition and excitation in dis-
tributed neural networks. Acta Psychologica, 101, 159Ð178.
Korzyukov, O., Winkler, I., Gumenyuk, V., Alho, K., &
Näätänen, R. (2003). Processing abstract auditory features
in the human auditory cortex. Neuroimage, 20, 2245Ð2258.
Massaro, D. W. (1970). Perceptual processes and forgetting in
memory tasks. Psychological Review, 85, 411Ð417.
Mondor, T. A., & Zatorre, R. J. (1995). Shifting and focusing
auditory spatial attention. Journal of Experimental Psychol-
ogy: Human Perception and Performance, 21, 387Ð409.

19I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Morton, J., Crowder, R. G., & Prussin, H. A. (1971). Experi-
ments with the stimulus suffix effect. Journal of Experimen-
tal Psychology, 91, 169Ð190.
Näätänen, R. (1990). The role of attention in auditory informa-
tion processing as revealed by event-related potentials and
other brain measures of cognitive function. Behavioral and
Brain Sciences, 13, 201Ð288.
Näätänen, R., Lehtokoski, A., Lennes, M., Cheour-Luhtanen,
M., Huotilainen, M., Iivonen, A., Vainio, M., Alku, P., Ilmo-
niemi, R. J., Luuk, A., Allik, J., Sinkkonen, J., & Alho, K.
(1997). Language-specific phoneme representations re-
vealed by electric and magnetic brain responses. Nature,
385, 432Ð434.
Näätänen, R., Tervaniemi, M., Sussman, E., Paavilainen, P., &
Winkler, I. (2001). ‘Primitive intelligence’ in the auditory
cortex. Trends in Neurosciences, 24, 283Ð288.
Näätänen, R., & Winkler, I. (1999). The concept of auditory
stimulus representation in cognitive neuroscience. Psycho-
logical Bulletin, 125, 826Ð859.
Nairne, J. S. (2002). Remembering over the short-term: The
case against the standard model. Annual Review of Psychol-
ogy, 53, 53Ð81.
Opitz, B., Rinne, T., Mecklinger, A., von Cramon, D. Y., &
Schröger, E. (2002). Differential contribution of frontal and
temporal cortices to auditory change detection: fMRI and
ERP results. Neuroimage, 15, 167Ð174.
Paavilainen, P., Jaramillo, M., Näätänen, R., & Winkler, I.
(1999). Neuronal populations extracting invariant relation-
ships from acoustic variance. Neuroscience Letters, 265,
179Ð182.
Paavilainen, P., Simola, J., Jaramillo, M., Näätänen, R., & Win-
kler, I. (2001). Preattentive extraction of abstract feature
conjunctions from auditory stimulation as reflected by the
mismatch negativity (MMN). Psychophysiology, 38, 359Ð
365.
Picton, D. W., Alain, C., Otten, L., & Ritter, W. (2000). Mis-
match negativity: different water in the same river. Audiol-
ogy and Neuro-Otology, 5, 111Ð139.
Pisoni, D. B. (1973). Auditory and phonetic memory codes in
the discrimination of consonants and vowels. Perception &
Psychophysics, 13, 253Ð260.
Rinne, T., Antila, S., & Winkler, I. (2001). Mismatch negativity
is unaffected by top-down predictive information. NeuroRe-
port, 12, 2209Ð2213.
Ritter, W., Deacon, D., Gomes, H., Javitt, D. C., & Vaughan,
H. G. Jr (1995). The mismatch negativity of event-related
potentials as a probe of transient auditory memory: A re-
view. Ear and Hearing, 16,52Ð67.
Ritter, W., Gomes, H., Cowan, N., Sussman, E., & Vaughan, H.
G. Jr (1998). Reactivation of a dormant representation of an
auditory stimulus feature. Journal of Cognitive Neurosci-
ence, 10, 605Ð614.
Ritter, W., Sussman, E., Molholm, S., & Foxe, J. J. (2002).
Memory reactivation or reinstatement and the mismatch neg-
ativity. Psychophysiology, 39, 158Ð165.
Rorden, C., & Driver, J. (2001). Spatial deployment of attention
within and across hemifields in an auditory task. Experimen-
tal Brain Research, 137, 487Ð496.
Rovee-Collier, C., & Hayne, H. (1987). Reactivation of infant
memory: Implications for cognitive development. Advances
in Child Development & Behavior, 20, 185Ð238.
Saarinen, J., Paavilainen, P., Schröger, E., Tervaniemi, M., &
Näätänen, R. (1992). Representation of abstract attributes of
” 2005 Hogrefe & Huber Publishers Experimental Psychology 2005; Vol. 52(1):3Ð20
auditory stimuli in the human brain. NeuroReport, 3, 1149Ð
1151.
Scherg, M., Vajsar, J., & Picton, T. W. (1989). A source analysis
of the late human auditory evoked potentials. Journal of
Cognitive Neuroscience, 1, 336Ð355.
Sussman, E., Ritter, W., & Vaughan, H. G. Jr (1998). Predict-
ability of stimulus deviance and the mismatch negativity.
NeuroReport, 9, 4167Ð4170.
Sussman, E., Sheridan, K., Kreuzer, J., & Winkler, I. (2003a).
Representation of the standard: stimulus context effects on
the process generating the mismatch negativity component
of event-related brain potentials. Psychophysiology, 40,
465Ð471.
Sussman, E., Winkler, I., Huotilainen, M., Ritter, W., &
Näätänen, R. (2002). Top-down effects on stimulus-driven
auditory organization. Cognitive Brain Research, 13, 393Ð
405.
Sussman, E., Winkler, I., & Wang, W. J. (2003b). MMN and
attention: Competition for deviance detection. Psychophysi-
ology, 40, 430Ð435.
Teder-Salejärvi, W. A., Hillyard, S. A., Roder, B., & Neville, H.
J. (1999). Spatial attention to central and peripheral auditory
stimuli as indexed by event-related potentials. Cognitive
Brain Research, 8, 213Ð227.
Winkler, I. (1996). Necessary and sufficient conditions for the
elicitation of the mismatch negativity. In C. Ogura, Y.
Koga, & M. Shimokochi (Eds.) Recent Advances in Event-
related Brain Potential Research (pp. 36Ð44). Amsterdam:
Elsevier.
Winkler, I. (2003). Change detection in complex auditory envi-
ronment: Beyond the oddball paradigm. In J. Polich (Ed.),
Detection of Change: Event-Related Potential and fMRI
Findings (pp. 61Ð81). Boston: Kluwer Academic Publish-
ers.
Winkler, I., Cowan, N., Cse
´
pe, V., Czigler, I., & Näätänen, R.
(1996). Interactions between transient and long-term audi-
tory memory as reflected by the mismatch negativity. Jour-
nal of Cognitive Neuroscience, 8, 403Ð415.
Winkler, I., & Czigler, I. (1998). Mismatch negativity: deviance
detection or the maintenance of the “standard.”. NeuroRe-
port, 9, 3809Ð3813.
Winkler, I., Karmos, G., & Näätänen, R. (1996b). Adaptive
modeling of the unattended acoustic environment reflected
in the mismatch negativity event-related potential. Brain Re-
search, 742, 239Ð252.
Winkler, I., Korzykov, O., Gumenyuk, V., Cowan, N., Linken-
kaer-Hansen, K., Alho, K., Ilmoniemi, R. J., & Näätänen, R.
(2002). Temporary and longer retention of acoustic informa-
tion. Psychophysiology, 39, 530Ð534.
Winkler, I., Lehtokoski, A., Alku, P., Vainio, M., Czigler, I.,
Cse
´
pe, V., Aaltonen, O., Raimo, I., Alho, K., Lang, A. H.,
Iivonen, A., & Näätänen, R. (1999). Pre-attentive detection
of vowel contrasts utilizes both phonetic and auditory mem-
ory representations. Cognitive Brain Research, 7, 357Ð369.
Winkler, I., Schröger, E., & Cowan, N. (2001). The role of
large-scale perceptual organization in the mismatch negativ-
ity event-related brain potential. Journal of Cognitive Neuro-
science, 13, 59Ð
71.
Winkler, I., Sussman, E., Tervaniemi, M., Ritter, W., Horva
´
th,
J., & Näätänen, R. (2003). Pre-attentive auditory context ef-
fects. Cognitive, Affective, & Behavioral Neuroscience, 3,
57Ð77.

20 I. Winkler & N. Cowan: From Sensory to Long-Term Memory
Received October 15, 2003
Revision received December 5, 2003
Accepted December 15, 2003
Adress for correspondence
Istva
´
n Winkler
Institute for Psychology
Hungarian Academy of Sciences
P.O. Box 398
H-1394 Budapest
Hungary
Tel. +36 1 354 2296
Fax +36 1 354 2416
E-mail winkler@cogpsyphy.hu
Experimental Psychology 2005; Vol. 52(1):3Ð20 ” 2005 Hogrefe & Huber Publishers
